Cómo se dividen las células: bases de mitosis y meiosis
¿Cómo logra una célula dividirse sin perder información en el proceso? ¿Y por qué, en otros casos, esa división no solo conserva el ADN, sino que además genera diversidad genética? Estas preguntas están en el corazón de la biología celular y conectan directamente con fenómenos tan distintos como el crecimiento de un tejido, la cicatrización de una herida o la reproducción sexual.
La división celular no es un simple “partirse en dos”, sino un proceso altamente regulado en el que la célula coordina la replicación, organización y distribución precisa de su material genético. En este recorrido exploraremos los mecanismos fundamentales que lo hacen posible, desde el ciclo celular que prepara a la célula, hasta las estrategias especializadas de la mitosis, la meiosis y la fisión binaria.
Más que una secuencia de fases a memorizar, estos procesos constituyen sistemas dinámicos con lógica molecular propia. Comprenderlos permite interpretar cómo se mantiene la estabilidad genética en los organismos y, al mismo tiempo, cómo surge la variabilidad que sustenta la evolución.
1. El Ciclo Celular: Recapitulación
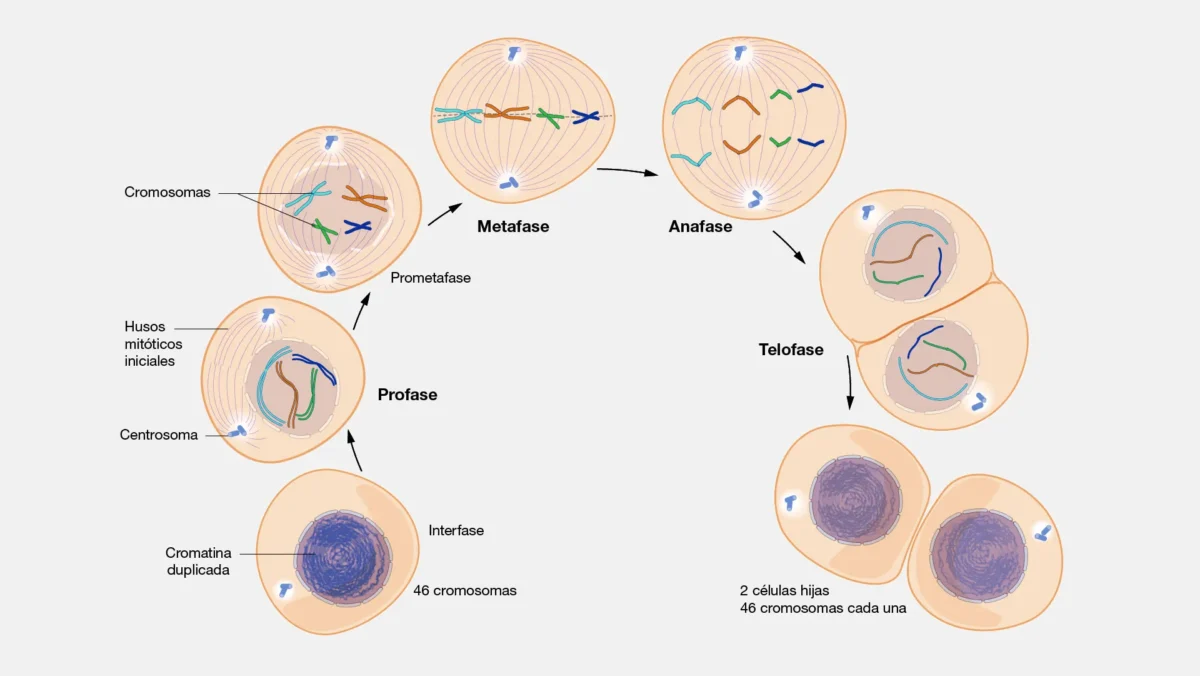
El ciclo celular, abordado en detalle anteriormente, comprende dos grandes etapas: la interfase (G1, S y G2) y la fase mitótica (M). Durante la interfase la célula crece, replica su ADN en la fase S —generando cromátidas hermanas unidas por cohesinas a lo largo de toda su longitud— y verifica la integridad del genoma en los puntos de control G1/S y G2/M. El resultado de la fase M es la producción de dos células hijas genéticamente idénticas a la progenitora. Los conceptos clave que se retomarán a lo largo de este tema son: la distinción entre centrómero (región del ADN) y cinetocoro (estructura proteica ensamblada sobre él), el papel de las cohesinas en la cohesión de cromátidas, y el rol del punto de control del huso (SAC) en metafase, que se desarrolla en detalle en las secciones siguientes.
2. La Mitosis: Segregación Genética
La mitosis (o cariocinesis, término que designa específicamente la división del núcleo, en contraste con la citocinesis o división del citoplasma) es un proceso continuo y altamente regulado que asegura que cada célula hija reciba un juego completo e idéntico de cromosomas. En los organismos pluricelulares, este mecanismo es esencial para el crecimiento, el desarrollo embrionario y la reparación de tejidos. Convencionalmente se divide en cinco etapas:
Profase
Las fibras de cromatina se condensan progresivamente gracias a la acción de proteínas llamadas condensinas, haciéndose visibles como estructuras definidas. Cada cromosoma duplicado está constituido por dos cromátidas hermanas unidas por el complejo de cohesinas a lo largo de toda su longitud. El nucléolo desaparece como consecuencia de la interrupción de la transcripción de genes de ARNr, cuya expresión activa depende de una cromatina descondensada. Los centrosomas migran hacia polos opuestos e inician la organización del huso mitótico mediante la polimerización de microtúbulos.
Prometafase
La envoltura nuclear se desintegra en pequeñas vesículas debido a la fosforilación de las láminas nucleares por CDK1. Los microtúbulos del huso invaden el espacio nuclear y se adhieren a los cinetocoros: estructuras proteicas multicomponente ensambladas sobre el ADN centromérico de cada cromátida. Los microtúbulos cinetocóricos establecen un proceso dinámico de «búsqueda y captura», con uniones inicialmente laterales que maduran a uniones terminales en el cinetocoro. En esta etapa comienza a operar el punto de control del ensamblaje del huso (SAC, del inglés Spindle Assembly Checkpoint), que detecta cinetocoros sin tensión o sin unión correcta, inhibiendo la progresión hasta que todos los cromosomas estén bialorientados.
Metafase
Los cromosomas, en su máxima condensación, se alinean en la placa ecuatorial (plano equidistante entre los dos polos). Cada cinetocoro queda unido a microtúbulos provenientes de polos opuestos (orientación bipolar o anfitélica). El SAC verifica que la tensión mecánica sobre los pares de cinetocoros sea simétrica; las uniones sin tensión adecuada (sintélicas, monotélicas o merotélicas) activan la inhibición del ciclo. Dado que en esta fase los cromosomas están en su máxima condensación y son más fácilmente individualizables, la metafase es la etapa preferida para el cariotipado clínico.
Anafase
La entrada en anafase está regulada por el Complejo Promotor de la Anafase/Ciclosoma (APC/C), ubiquitina-ligasa que marca para degradación a la securina, el inhibidor de la separasa. Una vez activada, la separasa corta el complejo cohesina que mantiene unidas a las cromátidas hermanas, permitiendo su separación. La unión en la región del centrómero es la última en romperse, ya que permanece protegida hasta este momento por mecanismos específicos de control. Se distinguen dos componentes:
- Anafase A: las cromátidas (ahora cromosomas hijos independientes) se desplazan hacia los polos por acortamiento de los microtúbulos cinetocóricos.
- Anafase B: los microtúbulos polares se elongan y deslizan entre sí gracias a motores moleculares (cinesinas), separando aún más los polos y alargando el huso.
Telofase
Los cromosomas alcanzan los polos y comienzan a descondensarse. Se reconstituyen las envolturas nucleares a partir de fragmentos de membrana, se despolimeriza el huso mitótico y el nucléolo reaparece a medida que se restablece la transcripción de ARNr. La célula retorna gradualmente al estado de interfase.
Citocinesis: División del Citoplasma
La fase mitótica culmina con la citocinesis, proceso que suele solaparse con las etapas finales de la mitosis. El mecanismo difiere según el tipo celular:
- Células animales: se ensambla un anillo contráctil de actina y miosina bajo la membrana plasmática, en el plano ecuatorial. Su contracción genera el surco de escisión que estrangula la célula hasta separarla en dos.
- Células vegetales: la pared celular rígida impide la constricción. En su lugar, vesículas del aparato de Golgi se fusionan en el plano ecuatorial formando el fragmoplasto y posteriormente la placa celular, que crece centrífugamente hasta fusionarse con la pared lateral existente.
3. La Meiosis: Especialización para la Reproducción Sexual
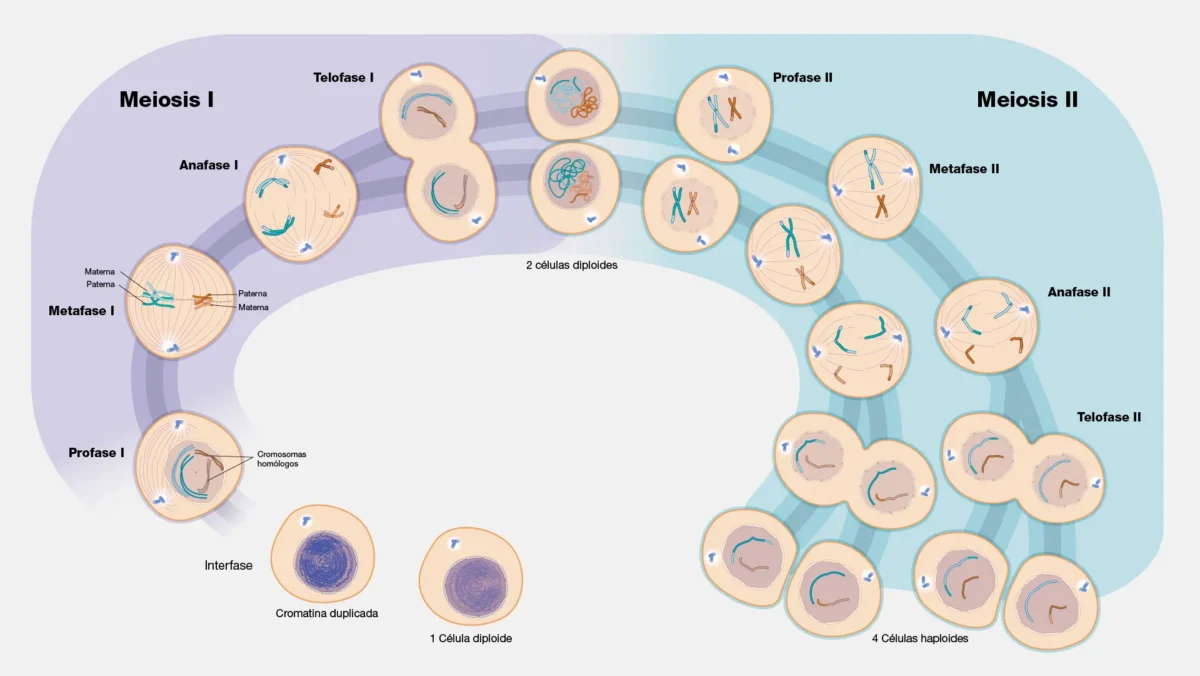
A diferencia de la mitosis, la meiosis es una división reductiva que en los animales produce gametos (óvulos y espermatozoides), y en plantas y hongos produce esporas a partir de las cuales se desarrollarán los gametofitos (que a su vez generarán gametos por mitosis). El resultado es células haploides (n), con la mitad del número de cromosomas de la célula progenitora diploide. La clave de esta reducción es que la meiosis consiste en una sola ronda de replicación del ADN seguida de dos divisiones nucleares consecutivas: precisamente porque la segunda división no va precedida de una nueva replicación, el número cromosómico se reduce a la mitad.
Meiosis I (División Reduccional)
Profase I
Es la fase más larga y compleja de toda la meiosis, con eventos sin equivalente en la mitosis. Se subdivide en cinco estadios:
- Leptoteno: los cromosomas comienzan a condensarse y se hacen visibles. Las puntas de los cromosomas se anclan a la envoltura nuclear en estructuras llamadas placas de anclaje, facilitando la búsqueda del homólogo.
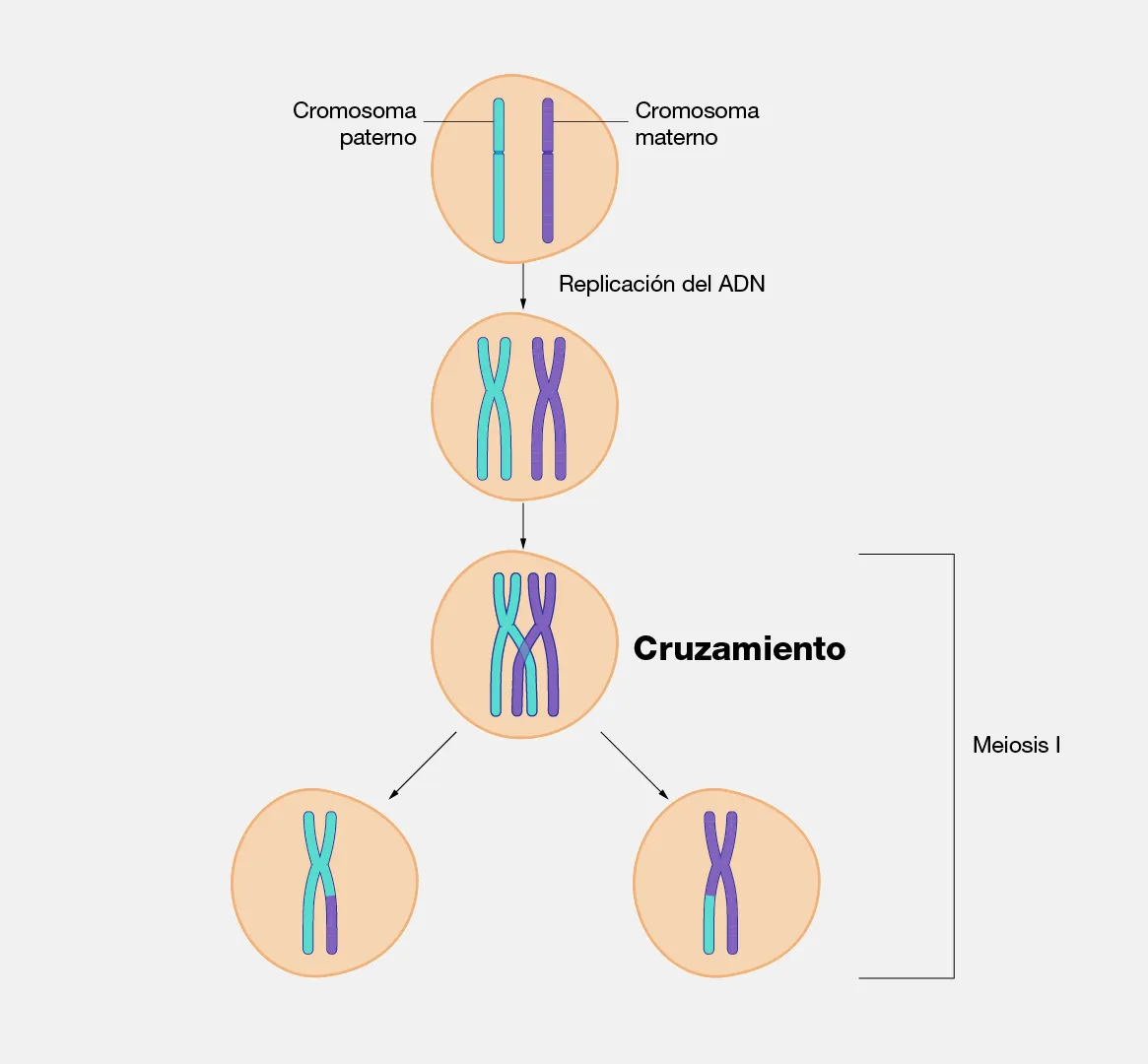
- Cigoteno: los cromosomas homólogos se aparean estrechamente mediante la formación del complejo sinaptonémico, una estructura proteica tripartita que los mantiene alineados con precisión a lo largo de toda su longitud, constituyendo bivalentes (o tétradas, formadas por cuatro cromátidas).
- Paquiteno: con el apareamiento completo de los cromosomas homólogos, ocurren intercambios físicos de segmentos de ADN conocidos como entrecruzamiento. Este proceso comienza con cortes controlados en ambas hebras del ADN, que luego son reparados mediante recombinación homóloga. Como resultado, se generan cromátidas con nuevas combinaciones de alelos, incrementando la variabilidad genética.
- Diploteno: el complejo sinaptonémico se disuelve y los homólogos comienzan a separarse, pero permanecen unidos en los puntos de sobrecruzamiento, ahora visibles como quiasmas. Los quiasmas no son meramente señales visibles de la recombinación ocurrida: cumplen una función mecánica activa, manteniendo la cohesión entre homólogos y siendo indispensables para su correcta segregación en anafase I. En ovocitos de mamíferos, la profase I puede quedar arrestada en diploteno durante años, hasta la ovulación.
- Diacinesis: los cromosomas alcanzan su máxima condensación, los quiasmas se desplazan hacia los extremos (terminalización), el nucléolo desaparece, la envoltura nuclear se fragmenta y se organiza el huso meiótico.
Metafase I
Los bivalentes (pares de cromosomas homólogos) se alinean en la placa ecuatorial. La orientación de cada bivalente es independiente, lo que genera variabilidad por segregación independiente (ley de Mendel). El SAC opera también en esta etapa, verificando la tensión sobre los cinetocoros.
Anafase I
Se separan los cromosomas homólogos entre sí (no las cromátidas hermanas). La unión a lo largo de los brazos cromosómicos se rompe, pero la región del centrómero permanece cohesionada, lo que permite que las cromátidas hermanas continúen unidas hasta la meiosis II.
Telofase I y Citocinesis
Se forman dos células, cada una con un juego haploide de cromosomas aún constituidos por dos cromátidas hermanas. Dependiendo de la especie, puede haber o no una interfase breve (sin replicación de ADN) antes de la meiosis II.
Meiosis II (División Ecuacional)
Esta fase es análoga a una mitosis, aunque opera sobre células haploides. La separasa, activada por el APC/C, cliva las cohesinas centroméricas restantes, separando las cromátidas hermanas. El resultado final son cuatro células haploides. El grado de variabilidad genética entre ellas depende del número y la posición de los sobrecruzamientos ocurridos en profase I y de la segregación independiente de los bivalentes en metafase I. En ausencia de sobrecruzamiento (como ocurre en machos de Drosophila melanogaster o en ciertas regiones del genoma), dos de las cuatro células serían genéticamente idénticas entre sí, y las otras dos también, aunque el par A sería diferente del par B.
4. Comparación entre Mitosis y Meiosis
| Característica | Mitosis | Meiosis |
|---|---|---|
| Propósito | Crecimiento, reparación tisular y reproducción asexual (en organismos que la utilizan como mecanismo reproductivo). | Producción de gametos (en animales) o esporas (en plantas y hongos) para la reproducción sexual. |
| Nº de divisiones | Una sola división nuclear y citoplasmática. | Dos divisiones nucleares y citoplasmáticas sucesivas, con una sola ronda previa de replicación del ADN. |
| Células hijas | Dos células hijas diploides (2n), genéticamente idénticas a la célula madre. | Cuatro células hijas haploides (n), genéticamente variables entre sí (según el número de sobrecruzamientos ocurridos). |
| Apareamiento de homólogos | No ocurre apareamiento entre cromosomas homólogos. | Ocurre sinapsis y sobrecruzamiento (crossing-over) durante la Profase I, mediados por el complejo sinaptonémico. |
| Comportamiento en Anafase | Se separan las cromátidas hermanas (mediante clivaje de cohesinas por la separasa). | Anafase I: se separan los cromosomas homólogos. Anafase II: se separan las cromátidas hermanas. |
| Puntos de control | G1/S, G2/M y SAC en metafase. | SAC activo en metafase I y metafase II, con regulación análoga adaptada a la dinámica meiótica. |
En síntesis: la mitosis preserva la identidad genómica de las células somáticas a lo largo del desarrollo y la reparación tisular, mientras que la meiosis introduce variabilidad genética (por recombinación y segregación independiente) y garantiza que la fusión de gametos en la fertilización restaure el número diploide característico de la especie.
5. Fisión Binaria y su Relación con la División Eucarionte
La fisión binaria es el mecanismo de división celular de los organismos procariontes (bacterias y arqueas). Es un proceso continuo más rápido que la división eucarionte, adaptado a genomas más compactos y a la ausencia de compartimentos membranosos internos.
Características principales
El material genético procarionte consiste típicamente en un único cromosoma circular de ADN, aunque muchas bacterias poseen además plásmidos. Al no existir núcleo verdadero, no ocurre cariocinesis ni envoltura nuclear que desintegrar y reconstituir.
La segregación de los cromosomas en procariotas no ocurre de forma pasiva. Existen sistemas activos que dirigen las copias del ADN hacia los polos de la célula. Algunos de estos mecanismos utilizan filamentos proteicos que se ensamblan entre las copias del cromosoma y generan fuerzas de empuje, funcionalmente análogas a las que ejercen los microtúbulos en células eucariotas.
La división del citoplasma en procariotas está mediada por FtsZ, que forma un anillo dinámico (Z-ring) en el plano de división y recluta los componentes del divisoma responsables de sintetizar el septo. FtsZ y la tubulina eucarionte son homólogos estructurales: comparten un plegamiento tridimensional similar y la capacidad de polimerizar de forma dependiente de GTP, aunque presentan baja identidad de secuencia. Esta relación indica que ambas proteínas descienden de un ancestro común y sugiere que elementos del citoesqueleto eucarionte tienen un origen evolutivo en proteínas ancestrales de procariotas.
Conclusión
Los procesos de división celular descritos en este tema —mitosis, meiosis y fisión binaria— representan tres soluciones evolutivas al problema fundamental de transmitir información genética de forma fiel y, cuando es adaptativamente ventajoso, con variabilidad. Lejos de ser procesos independientes, están unidos por una lógica molecular común: la necesidad de replicar el ADN con precisión, segregar las copias de forma equitativa y coordinar la división nuclear con la citoplasmática.
La mitosis encarna el principio de fidelidad: su entramado de puntos de control (G1/S, G2/M y SAC) y su maquinaria de cohesinas, separasa y APC/C garantizan que cada célula hija reciba exactamente el mismo complemento cromosómico que la progenitora. Cualquier fallo en esta fidelidad tiene consecuencias directas sobre la viabilidad celular y, en el contexto del organismo, puede desencadenar procesos tumorales.
La meiosis incorpora deliberadamente la variabilidad como función: la recombinación en profase I, y la segregación independiente de bivalentes en metafase I, generan una diversidad alélica que es el sustrato sobre el que opera la selección natural. Al mismo tiempo, la reducción del número cromosómico garantiza que la fertilización restaure la ploidía de la especie sin duplicarla generación tras generación.
En este sentido, mitosis y meiosis no son simplemente variantes de un mismo proceso: son estrategias complementarias que sostienen la estabilidad del genoma dentro de un organismo y la diversidad genética entre organismos.
La fisión binaria, aunque más simple en apariencia, comparte con la mitosis principios homólogos a nivel molecular que revelan una continuidad evolutiva profunda entre procariontes y eucariontes. Su estudio no es solo un ejercicio comparativo: la fisión binaria es la base de la proliferación bacteriana y, por tanto, de fenómenos con enorme relevancia biomédica como la resistencia a antibióticos y la dinámica de las infecciones.
En conjunto, comprender estos tres procesos con rigor mecanicista —no como secuencias de pasos a memorizar, sino como sistemas regulados con lógica causal— proporciona la base conceptual para abordar temas posteriores como la genética mendeliana, la oncología molecular, la biología del desarrollo y la evolución.