Ciclo Celular: Fases, Control y Claves para Comprender la División Celular
Introducción
El ciclo celular se define como una serie ordenada de eventos que involucran el crecimiento y la división celular, culminando en la producción de dos nuevas células hijas genéticamente idénticas. Este proceso es fundamental para la vida, permitiendo el desarrollo de organismos multicelulares a partir de un cigoto unicelular, así como la renovación y reparación de tejidos dañados.
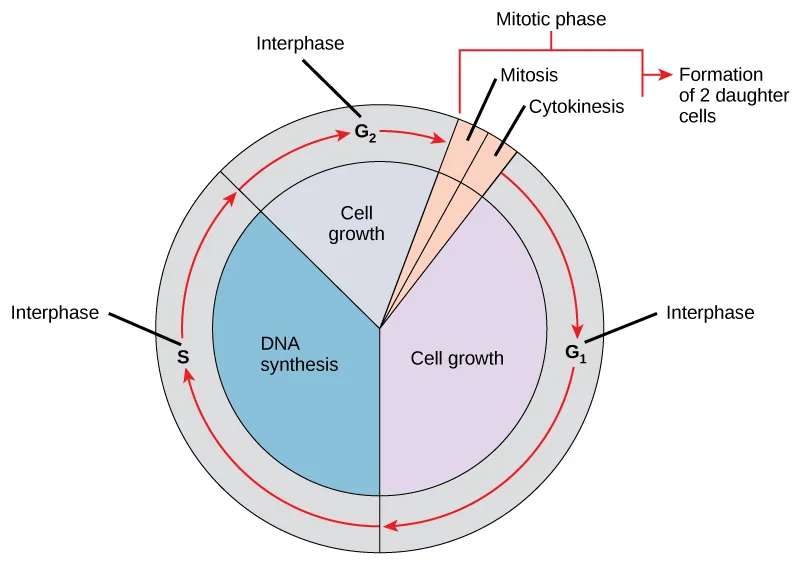
El ciclo se divide en dos fases principales: la interfase, donde la célula crece y replica su ADN, y la fase mitótica (fase M), donde ocurre la segregación del material genético y la división del citoplasma. La duración del ciclo es altamente variable; mientras que algunas células humanas en cultivo completan el proceso en aproximadamente 24 horas, las células embrionarias de organismos como Drosophila melanogaster o Xenopus laevis pueden hacerlo en pocos minutos.
Etapas del Ciclo Celular
La mayor parte de la vida de una célula transcurre en la interfase, que comprende aproximadamente el 90–95 % de la duración total del ciclo. Aunque morfológicamente el núcleo parece uniforme bajo el microscopio, a nivel molecular es una etapa de intensa actividad bioquímica. Esta se subdivide en tres etapas:
Fase G1 — Primer intervalo
Es un periodo de intenso crecimiento y actividad metabólica tras la mitosis, y suele ser la fase más larga y variable. La célula recupera su actividad biosintética, incrementando la síntesis de proteínas y duplicando orgánulos como mitocondrias y ribosomas, hasta alcanzar un tamaño óptimo. Se acumulan nucleótidos, proteínas y reservas de energía necesarias para la replicación del ADN. En esta fase también se integran señales extracelulares como factores de crecimiento.
Aquí se localiza el punto de restricción (en células de mamífero) o START (en levaduras como Saccharomyces cerevisiae), considerado el «punto de no retorno» inicial, donde la célula evalúa condiciones internas (tamaño, daño en el ADN) y externas (nutrientes, señales mitogénicas).
Si las condiciones no son favorables o la célula se especializa funcionalmente, puede salir del ciclo y entrar en un estado denominado fase G0. En este estado, la célula no se divide, pero permanece metabólicamente activa, llevando a cabo funciones especializada. Este estado puede ser reversible (quiescencia, como en linfocitos en reposo) o irreversible (estado postmitótico, como en neuronas y cardiomiocitos maduros).
Fase S — Síntesis de ADN
En esta etapa ocurre la replicación del ADN de manera altamente regulada y semiconservativa. Para comprender correctamente los cambios en el contenido genético, es necesario distinguir dos parámetros independientes:
- Ploidía (n): número de conjuntos de cromosomas homólogos. Una célula somática humana es diploide (2n = 46) y esta condición no cambia durante el ciclo celular mitótico.
- Contenido de ADN (C): cantidad de ADN por dotación haploide de referencia. Una célula diploide en G1 tiene contenido 2C. Al duplicar su ADN en fase S, pasa a 4C, pero sigue siendo 2n.
Cada cromosoma queda constituido por dos cromátidas hermanas idénticas unidas por cohesinas. Las cohesinas son complejos proteicos de la familia SMC (Structural Maintenance of Chromosomes) que se cargan sobre el ADN durante la replicación. Su eliminación es un proceso regulado en dos etapas: remoción parcial en los brazos cromosómicos durante profase y escisión en los centrómeros por separasa durante la anafase, evento que desencadena la separación de cromátidas.
La síntesis de histonas aumenta significativamente para permitir el empaquetamiento del nuevo ADN en nucleosomas. Asimismo, se duplica el centrosoma.
Fase G2 — Segundo intervalo
Corresponde al periodo final de preparación antes de la mitosis. La célula repone sus reservas energéticas y sintetiza proteínas necesarias para la dinámica cromosómica. Además, se producen cambios estructurales importantes, incluyendo reorganización de orgánulos y preparación del huso mitótico.
Se activa un control estricto para verificar que la replicación del ADN se haya completado correctamente. Si se detectan daños o replicación incompleta, se detiene el ciclo para permitir su reparación.
A nivel molecular, el punto de control G2/M distingue dos señales: (a) replicación incompleta, detectada por la persistencia de RPA sobre ADN de cadena sencilla en horquillas paradas, que activa ATR; y (b) roturas de doble cadena (DSBs), detectadas por ATM. Ambas vías convergen en la inhibición de CDK1 mediante la inactivación de la fosfatasa Cdc25C, impidiendo la entrada en mitosis.
Fase M — Mitótica
La fase M comprende dos procesos secuenciales: la mitosis (cariocinesis) y la citocinesis. La mitosis se divide en cinco etapas morfológicamente reconocibles:
- Profase: Los cromosomas se condensan progresivamente mediante la acción de las condensinas, lo que permite su correcta manipulación durante la división. El centrosoma duplicado comienza a separarse y se inicia la nucleación de los microtúbulos que formarán el huso mitótico. Paralelamente, la fosforilación de las láminas nucleares por complejos ciclina–CDK1 contribuye a la desestabilización de la envoltura nuclear, preparando su desensamblaje.
- Metafase: Tras la desintegración de la envoltura nuclear, los microtúbulos del huso capturan los cromosomas mediante sus cinetocoros. La correcta unión anfitélica —con microtúbulos provenientes de polos opuestos— genera tensión mecánica sobre las cromátidas hermanas, condición esencial para la estabilidad de las uniones. Este proceso es monitorizado por el punto de control del huso (SAC), que impide la progresión a anafase hasta que todos los cromosomas están correctamente biorientados. Una vez alcanzada esta configuración, los cromosomas se alinean en la placa ecuatorial bajo tensión bipolar, estableciendo la metafase propiamente dicha.
- Anafase: la activación de APC/C–Cdc20 degrada la securina, liberando a la separasa para escindir la cohesina centromérica. Anafase A: las cromátidas migran hacia los polos por acortamiento de microtúbulos cinetocóricos. Anafase B: separación adicional de polos.
- Telofase: los cromosomas llegan a los polos, se descondensa la cromatina, se reconstituye el envoltorio nuclear y desaparece el huso.
Finalmente, la citocinesis separa físicamente el citoplasma mediante la contracción de un anillo de actomiosina (en células animales) o la formación del fragmoplasto (en plantas).
Principios de Control del Ciclo Celular
El ciclo celular eucariota está regulado por mecanismos de vigilancia conocidos como puntos de control (checkpoints), que funcionan como un sistema de control de calidad capaz de detener la progresión del ciclo hasta asegurar que los procesos previos se han completado correctamente.
1. Punto de control G1 / Restricción
Es el principal punto de decisión y compromiso con la división celular. Evalúa la disponibilidad de nutrientes, el tamaño celular, las señales de crecimiento y, de manera crítica, la integridad del ADN. Si no se cumplen las condiciones, la célula puede detener el ciclo para reparación o desviarse hacia G0.
Molecularmente, el daño al ADN activa la vía ATM/ATR → CHK1/CHK2 → p53 → p21 → inhibición de CDK2/4.
2. Punto de control G2/M
Impide la entrada a la mitosis si la replicación del ADN no se ha completado o si existen daños en el material genético. Sus dos modos de activación son: (a) roturas de doble cadena activan ATM → CHK2 → inactivación de Cdc25C; (b) horquillas de replicación paradas activan ATR → CHK1 → inactivación de Cdc25C. En ambos casos, CDK1 permanece hiperfosforilado (inactivo) y no se inicia la mitosis.
3. Punto de control del huso — SAC (Spindle Assembly Checkpoint)
Ocurre durante la metafase y asegura la correcta unión anfitélica de todas las cromátidas hermanas a los microtúbulos del huso a través de sus cinetocoros. El sensor es el cinetocoro libre (sin microtúbulo o sin tensión mecánica), que cataliza el ensamblaje del MCC (Mitotic Checkpoint Complex: Mad2, BubR1, Bub3, Cdc20). El MCC inhibe APC/C–Cdc20, manteniendo estables la securina y la ciclina B.
Dado que la anafase es irreversible, la célula no progresa hasta que todos los cromosomas estén correctamente alineados, evitando errores de segregación como la aneuploidía.
La siguiente tabla resume los sensores y efectores moleculares de cada checkpoint:
| Checkpoint | Sensor principal | Efector / Mecanismo |
|---|---|---|
| G1 / START | ATM/ATR → CHK1/CHK2 | p53 → p21 → inhibe CDK2/4; degradación de Cdc25A |
| G2/M | ATM → CHK2 (DSBs); ATR → CHK1 (horquillas paradas) | Cdc25C inactivada → CDK1 hiperfosforilado → no entrada en M |
| SAC (huso) | Cinetocoros sin tensión → MCC activo | MCC inhibe APC/C–Cdc20 → securina y ciclina B estables → no anafase |
Regulación Molecular: Ciclinas, CDKs y sus Moduladores
La regulación molecular de los puntos de control depende de las ciclinas y las quinasas dependientes de ciclinas (CDKs), que actúan como reguladores positivos del ciclo celular mediante un sistema de interruptores moleculares (switches).
Las CDKs son quinasas de serina/treonina que fosforilan proteínas diana implicadas en procesos clave. Sin embargo, requieren la unión a ciclinas para activarse, formando complejos heterodiméricos funcionales. Los niveles de CDK1 y CDK2 permanecen relativamente constantes durante el ciclo, mientras que las ciclinas presentan oscilaciones cíclicas controladas por síntesis transcripcional y degradación mediante el sistema ubiquitina-proteasoma, lo que asegura la progresión unidireccional del ciclo.
La siguiente tabla resume los complejos ciclina–CDK más importantes y sus funciones:
| Complejo | Fase | Función principal |
|---|---|---|
| Ciclina D – CDK4/6 | G1 | Respuesta a mitógenos; fosforilación de Rb para liberar E2F |
| Ciclina E – CDK2 | G1 tardía / inicio S | Transición G1/S; inicio de la replicación |
| Ciclina A – CDK2 | S | Progresión de la replicación; inhibición de re-firing de orígenes |
| Ciclina A – CDK1 | G2 | Preparación para mitosis; fosforilación de láminas nucleares |
| Ciclina B – CDK1 (MPF) | M | Entrada en mitosis; condensación cromosómica, rotura del envoltorio nuclear |
La actividad de estos complejos está finamente regulada por múltiples mecanismos:
- Fosforilación activadora: CAK (CDK-Activating Kinase) fosforila el residuo Thr160/161 del dominio T-loop de las CDKs.
- Fosforilación inhibidora: la quinasa Wee1 (y Myt1 en membranas) fosforila Tyr15 y Thr14 de CDK1, manteniéndolo inactivo. La fosfatasa Cdc25 revierte esta inhibición.
- CKIs (Inhibidores de CDKs): p21 (CDKN1A), p27 (CDKN1B) y p16 (CDKN2A) inhiben diferentes complejos. p21 es diana transcripcional directa de p53.
El sistema APC/C y la unidireccionalidad del ciclo
El Complejo Promotor de Anáfase / Ciclosoma (APC/C) es una ubiquitina ligasa E3 que degrada selectivamente ciclinas y otros reguladores del ciclo. Su actividad depende de dos coactivadores:
- APC/C–Cdc20: activo durante la mitosis. Degrada ciclina B y securina, permitiendo la entrada en anafase.
- APC/C–Cdh1: activo en G1. Mantiene bajos los niveles de ciclinas S y M, estabilizando el estado G1 y previniendo una re-entrada prematura en S.
Reguladores negativos: p53 y Rb
La proteína p53, conocida como el «guardián del genoma», integra señales de daño genómico. En condiciones normales, p53 es mantenida a niveles bajos por MDM2 (una ubiquitina ligasa que la ubiquitina para su degradación proteasomal). Ante daño en el ADN, ATM/CHK2 fosforilan p53 e impiden su interacción con MDM2, permitiendo su acumulación. p53 activa la transcripción de p21, deteniendo el ciclo en G1 y G2. Si el daño es irreparable, p53 puede activar la apoptosis (vía PUMA, NOXA) o la senescencia.
La proteína Rb (retinoblastoma) regula la transición G1/S. En su forma hipofosforilada, Rb reprime a los factores de transcripción E2F, necesarios para la expresión de genes de fase S (ciclina E, ADN polimerasa α, timidina quinasa). El complejo ciclina D–CDK4/6 fosforila parcialmente Rb; el complejo ciclina E–CDK2 completa su hiperfosforilación, liberando E2F e iniciando el programa transcripcional de fase S. Esta liberación es el evento central del punto de restricción en mamíferos.
Conclusión
El ciclo celular constituye un sistema dinámico de control con propiedades que van más allá de una simple secuencia de fases: sus transiciones clave exhiben bistabilidad molecular, es decir, son interruptores (switches) con dos estados estables separados por umbrales de activación agudos. Esto se ha demostrado con mayor claridad en la transición G2/M, donde el bucle de retroalimentación positiva CDK1–Cdc25–Wee1 crea un sistema de todo-o-nada que hace que la entrada en mitosis sea abrupta e irreversible.
Esta arquitectura de control tiene consecuencias biológicas profundas: garantiza que la célula no quede atrapada en estados intermedios y que la información genética se transmita con fidelidad. Cuando estos mecanismos fallan —por mutación de p53, amplificación de ciclina D, pérdida de Rb o inactivación de checkpoints— la célula adquiere capacidad proliferativa descontrolada, uno de los hallmarks del cáncer.
Desde la perspectiva farmacológica, la comprensión de estos mecanismos ha generado una clase de fármacos dirigidos a componentes específicos del ciclo: los inhibidores de CDK4/6 (palbociclib, ribociclib, abemaciclib), aprobados para cáncer de mama, actúan bloqueando la fosforilación de Rb y deteniendo la progresión G1/S. El ciclo celular es, no solo un tema de biología básica, sino una de las dianas terapéuticas más activas en oncología molecular contemporánea.





